 Ferocactus cylindracious
hold the green chlorophyll necessary for photosynthesis; everything is armed with a fierce covering of thorns or spines, and nothing creeps along the baking ground. Cacti are iconic examples of this adaptation to a desert environment and a number of species are particular to the Sonoran desert of Arizona, and arid southwestern US/northwestern USA, including the barrel cacti (genus: Ferocactus; literally "ferocious cactus"!) a group of about 30 species that grow in a characteristic squat barrel-like shape. Their leaves have been modified into sharp curved hooks that grow in rows down ridges on the plant. Defense is particularly important in an environment where replacing lost resources is difficult, and in addition to their wicked hooks barrel cacti produce nectar from modified spines along the ridges to attract ants which provide extra defense against anything that may somehow get beyond the layer of spines. The ridges themselves allow the cactus to expand as it absorbs water when it rains or contract as it dries out in the longer periods between drinks. Interestingly (to me) the relationships and distinctness of species within this group are poorly understood, and it appears that some species currently thought to be Ferocactus may actually be more closely related to other cacti (Hernandez-Hernandez et. al., 2010). Even within Ferocactus morphology and genetics fail to conclusively establish a reliable family tree and telling species apart in the field can be difficult. I am not sure if what I thought were two species at the time were indeed different: Ferocactus wislizeni typically has orange/red flowers (above) and F. cylindracious has typically yellow flowers (at top) but both species can sometimes be found with the other color (is this convergent evolution of both flower colors or could it be species swapping genes and flower color by interbreeding?). Unsurprisingly for groups of plants that are hard to tell apart, molecular data suggests that the whole group are a very recent radiation, having all differentiated from a common ancestor not long ago (about 5 million years (Vazquez-Sanchez et. al., 2013)), giving little time for obvious reliable differences to occur between species. More intriguingly is that even in the short time I was there I saw several "species" growing in amongst each other; How species arise (grow apart from a common ancestor) in a short space of time and with limited spatial or environmental separation are questions that continue to fascinate and amuse many biologists, like myself.
Hernandez-Hernandez et. al., 2010; Phylogenetic relationships and evolution of growth form in Cactaceae (Caryophyllales, Eudicotyledoneae); American Journal of Botany, 98(1)
Vazquez-Sanchez et. al., 2013; Molecular phylogeny, origin and taxonomic implications of the tribe Cacteae (Cactaceae); Systematics and Biodiversity, 11(1)
0 Comments
 Cypripedium acaule (Pink Lady's Slipper Orchid) New York State, 2014 Maybe unsurprisingly the freezing months of winter have suppressed any happy thoughts of green herbage and summer flowers, but with Rochester experiencing OVER-freezing temperatures for the first time in living memory, it has been hard not to dream of thaw, the emergence of life again, and resurrect Tuesday Tracheophyte. Last year was my first North American spring and I was determined to hunt out some of the more iconic (but difficult to find) plants that are tucked away in small patches of remnant forest that have survived the largely cultivated and industrialized north-east. To have escaped attention these patches are generally poor land for farming and hard to access, often being rugged canyons or undrainable bogs and swamps. Happily there are a number of interesting plants that make a speciality living in the difficult conditions of being permanently waterlogged. One of the most spectacular of these groups are the slipper orchids (Cyprepedium spp.) with almost 60 species distributed across North and South America, Europe and Asia. There are several species locally, however the specificity of their growing requirements, fragmented habitat and poor reproductive success means many species are rare or endangered and difficult to find. Some have also become locally extinct through flower picking and over-zealous harvesting for the horticultural trade. Interestingly the pollination system of these plants is similar to that of the Australian orchid in my first post, although the two groups of species are not particularly closely related. Bees, attracted to the bright color and sweet scent of the flowers enter through a hole in the center of the pouch but on finding no nectar reward are unable to escape without brushing against the pollen sacks which are then hopefully transferred to the next flower. Individual bees apparently learn relatively quickly to avoid these flowers and successful pollination is less than 10%, however plants are also able to reproduce clonally (bulbs bud off) forming colonies. In fact so low is traditional reproduction in C. acaule that multiple successive years of fruiting reduces the health and vigor of plants in future years. Two locations in the Rochester area offer a decent chance of finding one or both of two species, C. acaule (Pink Lady's Slipper) and C. parviflorum (Yellow Lady's Slipper) although it is hard to find any exact details about where and when is the best time to go hunting . Our first expedition was to Bergen Swamp, a small reserve (and National Natural Landmark) managed by a local preservation society, biologically fascinating but effectively anonymous (whether this is deliberate or not, I am uncertain).
Thus buoyed I set out the next weekend to find a second species, this time in the affiliated, but distant, Zurich Swamp. Here again I was frustrated until the last few yards of my walk when I stumbled across a single pink flower of C. acaule. This species is particularly interesting as it inhabits highly acidic (pH 4-4.5) sphagnum moss beds. Such conditions are intolerable to most species, but the extreme chemistry actually inhibits the growth of fungi that would otherwise rot the bulbs of the orchids and they are unable to survive anywhere else.
The mosquitos were so thick that they can be seen in the photograph landing on the stem of the flower as I photograph it (and may explain my hasty and poor photography). This year I am resolved to return armed with military grade insect repellant and several layers of chain-mail or equivalent.  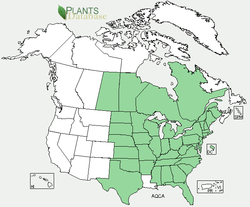 Aquilegia canadensis New York State 2014 From the name it seems reasonable that the Canada Columbine (ala 'canadensis') should at least be pretty much at home in the North of America, although it's alternative common name, the Eastern Red Columbine seems to confuse any obvious geographic affinities a little. In fact it appears that the Canada Columbine is perfectly at home on the majority of the North American continent, only prevented from extending entirely to the west by the Great Plains and/or Great Continental Divide. Being unable to cross these large unwooded expanses fits with a plant that prefers semi-shaded woodland fringes and wet river edges, and I have found patches of these attractive red flowers exploding out from the edges of swamps and tree-fall patches in denser forest (this picture was taken in Bergen Swamp, upstate NY). Sprays of vivid red flowers are hard to miss, and apparently humming birds find them hard to resist as well. In what appears to be becoming a theme, Columbines are a model system for the study of adaptation and evolution of switches in pollination syndrome between birds (red, with associated chemical pathways) and insects (blue, with alternate chemical pathways). Indeed, so remarkable are these contrasting colors in cultivated varieties that a favorite old Australian toffee brand, with red or blue wrappers, is named after them.
 Melaleuca aridus Simpson desert 2013 Following on from last week, this plant follows the theme of bird pollinated flowers from arid Western Australia. It is also a little controversial, and I have a conflict of interests. Until recently, the plant pictured above was generally considered to belong to the genus Calothamnus, closely related to another genus of shrubby plants, Melaleuca (below, right). For some time there was disagreement as to whether there was sufficient visible differences between these two groups to justify this separation. However using molecular sequence data for my honors project, I showed that as far as relationships go, these two groups of plants are mixed in together, and should be considered one and the same. Essentially Calothamnus are just slightly odd Melaleucas. Specifically, they have single flowers, rather than clusters, and these have squeezed their stamens together to form an elongated tube, the entrance to which is covered by a fringe of anthers (yellow capsules) holding the pollen. This is classic evolution toward specialized bird pollination, where the deep curved flower forces the bird to push it's bill well down into the nectaries, receiving a face-full of pollen for it's trouble. Compare this to the classical Melaleuca flower (below right) which has multiple flowers arranged in a cluster, each holding it's stamens much more openly, allowing access not only to birds but to insects and other animals as well. The whole cluster of flowers acting as a pollen brush. It is an interesting hypothesis that the evolution of more specialized bird pollinated flowers in the group-formerly-known-as-Calothamnus may be beneficial in moving pollen between the small, highly fragmented and widely dispersed populations of these plants.
The nuts (seed capsules) formed from fertilized flowers are incredibly hard and thick-walled, providing excellent protection from the harsh environment and predation. Apart from being strikingly elegant, the flowers of this particular species (Melaleuca aridus) emerge as pale peachy pink (above left) and mature to the vibrant red over a day or so. As with most Myrtaceae they have no perceptible smell. The plants themselves grow to be lanky scrubby shrubs roughly 2m high with round pencil like leaves. Similar to last weeks plant (Leptosema) these Melaleucas have specialized leaves that are an adaptation to highly arid conditions, being cylindrical to minimize surface area and water loss, with a layer of furry hairs to further help insulate the leaf. Conserving every drop of water is important to a plant that grows on ridges and rocky rises surrounded by deserts of deep dry sand, a problem faced by many Australian plants, which have evolved many varied and fascinating ways of adapting to meet the challenge.
Photos: R.D. Edwards  Leptosema chambersii
What happens if you are a small plant that is pollinated by birds or mammals rather than insects, but you have no branches for the animal to land on to access your flowers? Grow your flowers next to the ground so that they are easier to get to! Leptosema is a small genus of pea flowers native to semi-arid Australia that have done exactly this. Each small straggly plant produces bright red flowers around its base (red is more attractive to vertebrates than insects) where they are within easy reach of any animal on the ground. Flowers produce a rich nectar reward that is attractive to mammals and birds, and have been turned upside down so that as an animal pushes forward into the nectary it receives a brushing of pollen on the top of the head. This is subsequently transferred to the female parts (stigma - large curved rod structure seen here) of the next flower it visits. The tall narrow flowers inspired the name lepto (slender) sema (banner) from the Greek. Interestingly it appears that shifting from insect to bird pollination has evolutionary consequences, with reduced species diversity and potentially greater extinction risk: http://www.biomedcentral.com/1471-2148/14/43 Arid growing conditions have also led to the loss of true leaves (leaves loose water easily) in these species, and the development of flattened stems which hold chloroplasts and photosynthesize instead. Photo: R.D. Edwards  Pterostylis nutants (nodding greenhood)
A native Australian terrestrial orchid, widespread throughout eastern Australia, favoring moist protected gullies in schlerophyll (meaning "dry leaves") open eucalypt forest. Like many orchids, this plant has an interesting way of being pollinated (pollination syndrome). An insect is attracted to the flower (presumably by scent or pheromones) and enters under the tongue ("labellum", curving out mid right) seeking a (nonexistent) reward. The tongue closes behind the insect forcing it to crawl up into the translucent windows, towards the light and then eventually through the larger chamber back down and out (anti clockwise from top here). On the way they must pass the pollinea (pollen bundles, black blob, extreme left) that stick to its body. The insect then pushes through the beak of petals/sepals at the bottom, leaving unrewarded, annoyed, loaded down with pollen, but presumably not having learned anything as they will fall for it all over again, this time fertilizing the next flower with their pollen passenger. Photo: R.D. Edwards |
Bort Edwards







 RSS Feed
RSS Feed
